20. Модифицированная модель Лотки – Вольтерра и проблема обучаемости
Механизмы принятия административных решений в системе образования могут быть эффективными, если им предшествуют предварительные расчеты, основанные на математических моделях. Особенно остро в последнее время стоит вопрос о выявлении конкретной способности индивидуумов к обучению («обучаемости»), количественной мерой которой может служить время, необходимое для прочного усвоения определенного объема информации. Без решения задачи Обучаемости в зависимости от времени невозможно принятие научно обоснованных решений по распределению часов, отведенных на тот или иной предмет (блок предметов) в образовательных стандартах, по объему нагрузки на школьников и студентов, и т. п. Именно поэтому возрастает значимость тестирования, позволяющего делать по результатам обучения определенные «срезы знаний» во времени.
Тем не менее, важность исследования обучаемости в зависимости от времени в явной форме, в виде определенной функции ε (T) также несомненна. Здесь требуются глубокие психолого-педагогические подходы, информация о которых на сегодняшний день слишком мала.
В рамках данного учебника можно лишь продемонстрировать фундаментальность функции обучаемости, являющейся необходимым элементом деятельности не только в человеческом обществе. Особенно велика роль обучения в условиях соревнований, конкурентной борьбы, борьбы «за выживание», борьбы «на уничтожение» и т. п.
Ниже рассмотрим две такие системы с элементами обучаемости, которые базируются на расширении известной в синергетике и биологии модели Лотки – Вольтерра (модели «хищник - жертва»):
![]() = α1N – β1NM, (1)
= α1N – β1NM, (1)
![]() = – α 2 M + β2 NM,
= – α 2 M + β2 NM,
Где N – количество «кроликов – жертв» в системе с неограниченным резервом травяной пищи для них, а M – количество «рысей - хищников», рост которых зависит от вероятности встреч хищника с жертвой. Эта вероятность пропорциональна произведению NM. Одновременно, от вероятности такой встречи уменьшается число кроликов. Поэтому в верхнем уравнении системы (1) слагаемое в правой части, пропорциональное произведению NM, стоит со знаком минус. Коэффициенты α1, α2, β1 и β2 в этой системе являются константами, а их смысл обсуждается в любом издании (область биологии и синергетики).
Новая постановка задачи выживаемости (конкурентоспособности), которую предлагается проанализировать, заключается в том, чтобы ввести в систему уравнений (1) зависимость коэффициентов β1 и β2 от времени и считать их связанными с функцией обучаемости ε (T) в виде β1(T) = β2(T) =  , где εN (T) – обучаемость кроликов, А εM (T) – соответственно, обучаемость хищников во времени.
, где εN (T) – обучаемость кроликов, А εM (T) – соответственно, обучаемость хищников во времени.
Очевидно, что чем быстрее растет обучаемость кроликов по сравнению с обучаемостью хищников, тем меньше величина β1 (T) И β2 (T), что в свою очередь приводит к благоприятным условиям «выживания» кроликов. Если приблизиться к реальности, то εN (T) и εM (T) могут быть периодическими функциями с разными частотами, могут зависеть от N и M, что будет означать распределение кроликов и рысей по их способностям к обучению и т. п. В этих случаях, результаты расчетов для зависимости N(T) и M(T) заранее не предсказуемы даже качественно и решения будут зависеть от всех констант и параметров распределений.
Имеют право на существование и другие направления исследований в рамках модифицированной модели Лотки – Вольтерра с элементами обучаемости. Если говорить о борьбе за выживание среди людей, то в первую очередь, следует иметь в виду боевые действия. Так как время, деньги, ресурсы и информация являются всеобщими эквивалентами обмена, то и переменная T в системе уравнений не обязательно играет роль времени. Так, например, во время театра военных действий, величины ![]() и N(T) Могут иметь смысл скорости изменения (и численности) военнослужащих в зависимости от денежных затрат, выделенных ресурсов, снабжения информацией и т. п.
и N(T) Могут иметь смысл скорости изменения (и численности) военнослужащих в зависимости от денежных затрат, выделенных ресурсов, снабжения информацией и т. п.
Очевидно, что множества военнослужащих N и M являются одновременно и хищниками и жертвами, а систему уравнений может представить следующим образом:
![]() = α 1 (Np – N) – β1NM, (2)
= α 1 (Np – N) – β1NM, (2)
![]() = α 2 (Mp – M) - β2 NM.
= α 2 (Mp – M) - β2 NM.
Где Nр и Mр – константы, определяющие резервное количество военнослужащих, а β1 и β2 По прежнему могут быть связаны с функциями ε1(t) и ε2 (t), но уже не только от времени (хотя и это остается в силе), но и от денежных затрат, ресурсов и информации. Похожие уравнения можно ввести и в экономике, анализируя выживаемость фирм на рынке в условиях конкуренции. Но во всех рассматриваемых системах функция обучаемости (информированности, компетентности, профессионализма) является едва ли не основным фактором выживания.
Если, например, ε(T) = At + B, т. е. функция обучаемости имеет простейший вид линейной зависимости от времени, то коэффициенты А (выражающий среднюю способность индивидуумов к обучению) и B (определяющий стартовый уровень знаний, умений и навыков, и даже культуры) по величине, в общем случае, различны, что сказывается на конечном результате. Необходимо проведение практических исследований, прежде чем они могут быть использованы для решения задач в конкретной ситуации. По-видимому, А и B могут быть ранжированными параметрами, для которых необходимо вводить некоторый набор эталонов, как это делается, например, в психологии.
Тем не менее, сама система уравнений (модель) имеет прогностическую ценность, так как она позволяет, «играя с параметрами», выявить устойчивые решения и условия, при которых осуществляются различные «сценарии». В этом случае сценарии, определяемые уравнениями (1) и (2), могут служить основой для построения определенной деловой игры, в которой выигрышем является угадывание результатов решения, т. е. правильное прогнозирование поведения системы, развитие интуиции.
Приведем лишь два конкретных примера (а их много и обсуждать их следует отдельно) по результатам исследования модифицированной системы (1) при заданных параметрах функции обучаемости (рис.1а, б). Так как нелинейные уравнения не имеют общих аналитических способов их решения, то решение было получено на компьютере численными методами (программа «Mathcad»).
На рис.1а представлены изменения популяций хищников и кроликов в зависимости от времени (три первых диаграммы А, б,в и фазовая траектория (Г) для величин N и M) при εN(T )= Ct2+D и εM(T)= At+B, а на рис.1б при εN(T) = Ct + D и εM(T) = At+B.
Анализируя результаты расчетов, легко заметить, что вместе с увеличением количества кроликов во времени растет и число хищников, а фазовая диаграмма (Г) имеет вид разворачивающейся спирали. Если же проводить экспериментальные исследования, опираясь на информацию о числе хищников, то наблюдаемое увеличение их популяции могло бы навести на неверный вывод о том что, как следствие, количество кроликов должно уменьшаться, а оно также увеличивается (?!).
![]()
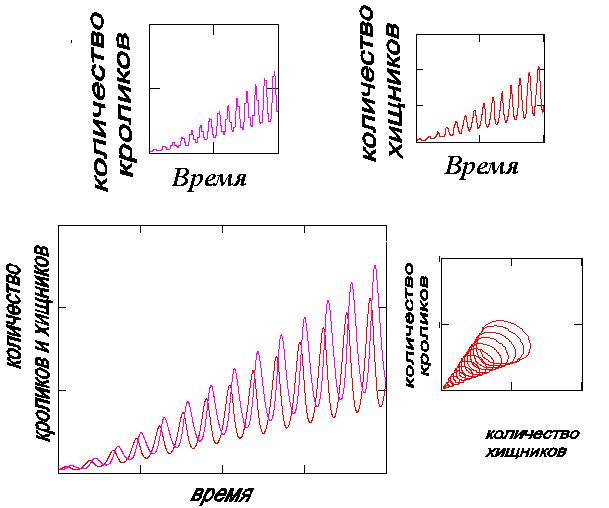
Рис. 1а. Результат решения уравнений (1) при нелинейной
зависимости εN(T) от времени.
![]()
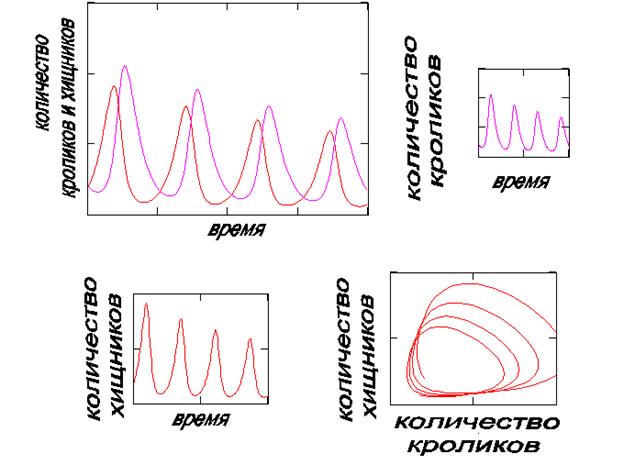
Рис. 1б. Результат решения уравнений (1) при линейной
зависимости εN(T) и εM(T) от времени (А > C)
В модельном эксперименте, представленном на рис. 1б, хищники обучаются немного быстрее, чем кролики и это приводит к постепенному уменьшению как жертв, так и их врагов. Интересным фактом является наличие периодичности во времени, похожей на «резонансные» движения качелей в первом случае (рис. 1а) и на затухающие колебания – во втором (рис. 1б). Очевидно, существует стационарный процесс саморегуляции количества хищников и кроликов, при котором совершаются колебания численности и тех и других во времени с постоянной амплитудой. Этот эксперимент уже поставлен природой. В Канаде была накоплена информация о количестве кроликов и рысей в лесу за несколько лет, которая позволила сделать вывод о стационарности процесса: колебания числа кроликов опережают колебания для количества рысей, о чем можно прочесть в учебной литературе по синергетике. Параметры β1 и β2 при стационарном состоянии биологической системы в модели (1) не зависят от времени, то есть являются постоянными. Математическое моделирование привело к адекватному описанию фактов.
При отсутствии у кроликов врагов (M = 0), решение верхнего уравнения в системе (1) приводит к экспоненциальной функции роста количества кроликов, что легко проверить самостоятельно. Известен факт резкого увеличения популяции кроликов в Австралии, где для них не нашлось естественных врагов.
Тем не менее, закон экспоненциального роста указывает лишь Тенденцию хода процесса, в то время как более детальное изучение закономерности роста кроликов (без врагов) показало, что этот рост следует по порядку числам в ряду последовательности Фибоначчи: 1,1,2,3,5,8,13,21,… , который строится по правилу: F(N+1) = F(N) + F(N –1), Где F(N) – число, стоящее в ряду последовательности на N-М месте. Этот экспериментальный факт показывает, что размножение кроликов происходит медленнее, чем при экспоненциальном росте.
| < Предыдущая | Следующая > |
|---|